Bibliographie
Ashmore, J. (2008). Cochlear outer hair cell motility. Physiological Reviews, 88, 173-210. https://doi.org/10.1152/physrev.00044.2006
Davis, A. C. (1989). The prevalence of hearing impairment and reported hearing disability among adults in Great Britain. International Journal of Epidemiology, 18 (4), 911-917. https://doi.org/10.1093/ije/18.4.911
Gilliver, M., Beach, E. F., & Williams, W. (2013). Noise with attitude : Influences on young people’s decisions to protect their hearing. International Journal of Audiology, 52 (sup1), S26-S32. https://doi.org/10.3109/14992027.2012.743049
Gilliver, M., Beach, E. F., & Williams, W. (2015). Changing beliefs about leisure noise: Using health promotion models to investigate young people’s engagement with, and attitudes towards, hearing health. International Journal of Audiology, 54 (4), 211-219. https://doi.org/10.3109/14992027.2014.978905
Hannula, S., Bloigu, R., Majamaa, K., Sorri, M., & Mäki-Torkko, E. (2011). Self-reported hearing problems among older adults: Prevalence and comparison to measured hearing impairment. Journal of the American Academy of Audiology, 22 (8), 550-559. https://doi.org/10.3766/jaaa.22.8.7
Kawase, T., Delgutte, B., & Liberman, M. C. (1993). Antimasking effects of the olivocochlear reflex. II. Enhancement of auditory-nerve response to masked tones. Journal of Neurophysiology, 70, 2533-2549. https://doi.org/10.1152/jn.1993.70.6.2533
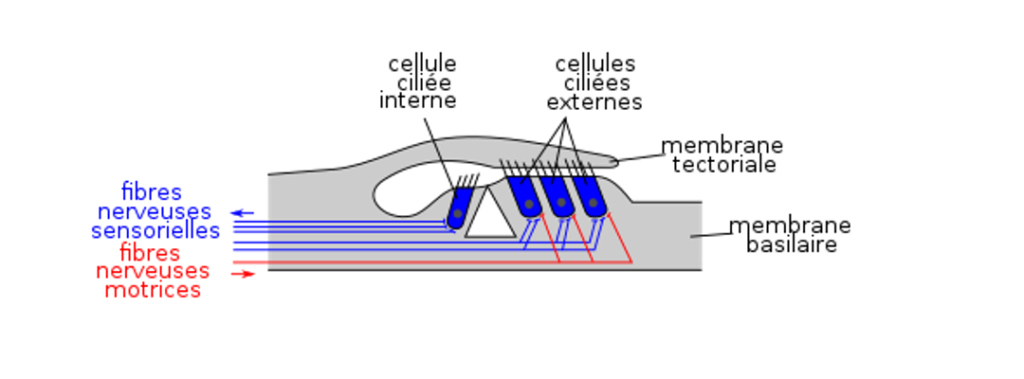
Liberman, M. C., & Kiang, N. Y. (1984). Single-neuron labeling and chronic pathology. IV. Stereocilia damage and alterations in rate and phase level functions. Hearing Research, 16, 75-90. https://doi.org/10.1016/0378-5955(84)90026-1
Pang, J., Beach, E. F., Gilliver, M., & Yeend, I. (2019). Adults who report difficulty hearing speech in noise: An exploration of experiences, impacts and coping strategies. International Journal of Audiology, 58 (12), 851-860. https://doi.org/10.1080/14992027.2019.1670363
Tremblay, K. L., Pinto, A., Fischer, M. E., Klein, B. E. K., Klein, R., Levy, S., Tweed, T. S., & Cruickshanks, K. J. (2015). Self-reported hearing difficulties among adults with normal audiograms: The Beaver Dam Offspring Study. Ear and Hearing, 36 (6), e290. https://doi.org/10.1097/AUD.0000000000000195
Zheng, Y., & Guan, J. (2018). Cochlear synaptopathy: A review of hidden hearing loss. Journal of Otorhinolaryngology Disorders & Treatment, 1 (1). https://dx.doi.org/10.16966/jodt.105


{kind=link}