Lorsque vous ne faites rien, que vous ne pensez à rien et que vous laissez votre cerveau se reposer, est-ce que votre cerveau « s’éteint » vraiment ?
En 1995, l’ingénieur biomédical Biswal a publié un article traitant de cette question. Il analysait des données d’imagerie par résonance magnétique fonctionnelle (fMRI) [1, 2]. Il essayait de supprimer le bruit causé par les mouvements associés à la respiration et au rythme cardiaque du signal représentant l’activité cérébrale réelle. Il a analysé la corrélation (c.-à-d. la relation) entre le signal IRMf dans le cortex moteur et d’autres régions du cerveau. À sa grande surprise, il découvrit que le cortex moteur était fonctionnellement connecté à d’autres régions motrices du cerveau, même lorsqu’une personne était au repos [2]. Ce type de « connexion » dite fonctionnelle signifie que lorsqu’une région du cerveau est activée ou engagée, l’autre région montre un schéma similaire d’activation ou d’engagement. Les régions peuvent ainsi être considérées comme « associées ». Cette connectivité (ou couplage) est connue sous le nom de connectivité fonctionnelle à l’état de repos (CFAR). Cette découverte a remis en question la notion bien établie selon laquelle l’activité cérébrale était principalement motivée par des tâches ou des stimuli externes. La découverte de la connectivité fonctionnelle à l’état de repos a ouvert de nouvelles possibilités pour étudier l’architecture fonctionnelle du cerveau.
En 2001, Raichle et ses collègues de la faculté de médecine de l’Université de Washington ont décrit l’activité des régions cérébrales qui étaient plus actives au repos comme représentant un état de base ou un « mode par défaut » du cerveau [3]. Ceci était basé sur l’observation répétée que les régions formant ce réseau présentaient une activité métabolique réduite lorsque les gens sont engagés dans des tâches cognitives – ces observations ont été effectuées à l’aide d’une technique appelée tomographie par émission de positrons (TEP), laquelle peut mesurer l’activité métabolique à l’aide d’une substance radioactive. En 2003, Greicius et ses collègues ont identifié des régions cérébrales avec une plus grande activité au repos et les ont nommées le réseau en mode par défaut (DMN). Les résultats étaient cohérents avec les travaux de Raichle et ses collègues appelés la ligne de base du « mode par défaut » [4].
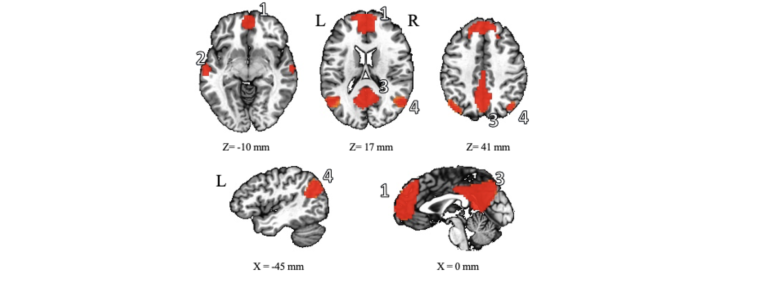
Le DMN présente une activité plus élevée lorsque le cerveau est au repos, plutôt qu’engagé dans des tâches cognitives, motrices ou langagières spécifiques, telles que la discrimination auditive et le traitement de la parole. Comme le montre la figure 1, ce réseau comprend les aires cérébrales suivantes : le cortex frontal orbital, le cortex préfrontal médial (MPFC), le cortex cingulaire antérieur (ACC), le cortex temporal latéral, le lobe pariétal inférieur, le cortex cingulaire postérieur (PCC), le cortex rétrosplénial et des structures telles que l’hippocampe et le gyrus parahippocampique. La robustesse du DMN à travers les études et la facilité avec laquelle il est observé ont contribué à stimuler l’acceptation et l’utilisation de l’IRMf à l’état de repos en tant qu’outil neuroscientifique rigoureux. Le nombre de publications portant sur la connectivité fonctionnelle au repos a décollé au milieu des années 2000 et des milliers d’articles sont maintenant publiés chaque année en utilisant cette technique. L’IRMf à l’état de repos est devenue un outil largement utilisé dans la recherche en neurosciences.
Figure 1. Le réseau en mode par défaut (DMN) chez l’humain, représenté ici en vue axiale (en haut) et sagittale (en bas), inclut : (1) le cortex préfrontal médian (MPFC)/cortex cingulaire antérieur (ACC), (2) le cortex temporal latéral, (3) le cortex cingulaire postérieur (PCC)/ cortex rétrosplénial, (4) le lobe pariétal inférieur, (5) l’hippocampe et le cortex [5, 6].
Bien que les fonctions précises du DMN ne soient pas encore entièrement comprises, la recherche suggère que ce réseau pourrait soutenir divers processus cognitifs autoréférentiels, comme l’autoréflexion, l’introspection, le voyage mental dans le temps (se souvenir du passé et envisager l’avenir) et la rêverie. De plus, plusieurs études ont montré que le DMN présente des anomalies dans de nombreux troubles neuropsychiatriques et neurologiques, comme la dépression, l’anxiété, l’autisme et la maladie d’Alzheimer, ce qui suggère que la connectivité fonctionnelle pourrait être utilisée en tant qu’indice de la santé du cerveau. Ainsi, la recherche sur le DMN recèle un potentiel important pour comprendre et traiter ces conditions.
En plus d’indexer la santé et la pathologie cérébrale, la connectivité fonctionnelle à l’état de repos peut indexer le vieillissement cérébral. En effet, de nombreuses études ont rapporté que la connectivité fonctionnelle est plus faible chez les personnes âgées par rapport aux personnes plus jeunes [7-10], mais les mécanismes sous-jacents n’ont pas encore été complètement élucidés. Il est bien établi que le cerveau adulte conserve la capacité de modifier sa structure et son fonctionnement à travers les expériences et l’apprentissage. Ce processus d’adaptation basé sur les expériences est appelé « plasticité cérébrale dépendante de l’expérience ». Ce type de plasticité cérébrale peut être associé à une amélioration des performances cognitives ou motrices. Par exemple, les chauffeurs de taxi développent une excellente mémoire spatiale ; à mesure que leur mémoire spatiale se développe, leur hippocampe augmente également [11].
Dans notre laboratoire, nous étudions l’impact du vieillissement et des expériences musicales sur la connectivité fonctionnelle. Nous recrutons des chanteurs, des instrumentistes et des non-musiciens adultes de tous âges pour étudier leur physiologie cérébrale au repos. Dans le cadre d’une expérience au repos, nous installons confortablement les participants dans l’appareil IRM et leur demandons d’ouvrir leurs yeux et de ne penser à rien de particulier, mais sans dormir. Les données IRMf sont mesurées pendant 10 minutes. En plus de mesurer la fonction cérébrale, nous mesurons également les performances cognitives et langagières, telles que la capacité d’attention auditive et le traitement de la parole, afin de déterminer s’il existe une relation entre l’activité cérébrale et la cognition, et si cette relation change avec l’âge et l’expérience musicale. Nous espérons que ce travail apportera de nouvelles lumières sur l’effet potentiellement transformateur des activités musicales et aidera à mettre en lumière le potentiel de ces activités pour atténuer ou ralentir le vieillissement cérébral et le déclin cognitif.
Références:
Biswal, B., et al., Functional connectivity in the motor cortex of resting human brain using echo-planar mri. Magnetic Resonance in Medicine, 1995. 34(4): p. 537-541.
Biswal, B.B., Resting state fMRI: a personal history. Neuroimage, 2012. 62(2): p. 938-44.
Raichle, M.E., et al., A default mode of brain function. Proceedings of the National Academy of Sciences, 2001. 98(2): p. 676-682.
Greicius, M.D., et al., Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proceedings of the National Academy of Sciences, 2003. 100(1): p. 253-258.
Raichle, M.E., The Brain’s Default Mode Network. Annual Review of Neuroscience, 2015. 38(1): p. 433-447.
Lu, H., et al., Rat brains also have a default mode network. Proceedings of the National Academy of Sciences, 2012. 109(10): p. 3979-3984.
Farras-Permanyer, L., et al., Age-related changes in resting-state functional connectivity in older adults. Neural Regeneration Research, 2019. 14(9): p. 1544-1555.
Ferreira, L.K. and G.F. Busatto, Resting-state functional connectivity in normal brain aging. Neuroscience & Biobehavioral Reviews, 2013. 37(3): p. 384-400.
Grady, C.L., O. Grigg, and C. Ng, Age differences in default and reward networks during processing of personally relevant information. Neuropsychologia, 2012. 50(7): p. 1682-1697.
Damoiseaux, J.S., et al., Reduced resting-state brain activity in the “default network” in normal aging. Cerebral Cortex, 2008. 18(8): p. 1856-1864.
Maguire, E.A., et al., Navigation-related structural change in the hippocampi of taxi drivers. Proceedings of the National Academy of Sciences, 2000. 97(8): p. 4398-4403.



{kind=link}